Mantophasmatodea
Mantophasmatodea é uma ordem de insetos descoberta no ano de 2002 pelo entomólogo alemão Oliver Zompro, do Instituto Max Planck de Limnologia. Foram apelidados de gladiadores, devido à "armadura" que cobre estes insetos no estágio de ninfa.
| Mantophasmatodea | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||
| Classificação científica | |||||||||||||
| |||||||||||||
| Famílias | |||||||||||||
| Austrophasmatidae | |||||||||||||
Suas relações filogenéticas mais próximas podem ser com Grylloblattodea e/ou Phasmatodea, mas a evidência morfológica é ambígua.
Habitam o oeste da África do Sul e Namíbia (maciço de Brandberg). Porém, fósseis do Eoceno sugerem uma distribuição mais ampla no passado.
A ordem foi descrita inicialmente a partir de cinco espécimes vivos encontrados na Namíbia e Tanzânia (Mantophasma zephyra e M. subsolana) e de um espécime fóssil do âmbar proveniente de uma coleção da University of Hamburg (Alemanha) e retirado do Mar Báltico, datado de 45 milhões de anos (Rhaptophasma kerneggeri).
História da Ordem editar

São indivíduos de difícil classificação, já que possuem uma mistura de características morfológicas de ordens diferentes. São facilmente confundidos com ninfas de louva-a-deus, apesar de não possuírem patas dianteiras raptoriais; diferem dos bichos-pau pelo hábito alimentar (são carnívoros, ao passo que os bichos-pau são herbívoros); não possuem apêndices especializados para pular como gafanhotos e grilos possuem, além de terem o órgão ovipositor e caracteres do tórax diferentes do que esses dois grupos.
Espécimes de Matophasmatodea já haviam sido coletados, montados e abrigados em coleções de museus há anos, porém eram classificados como ninfas de louva-a-deus. Em 1890, 5 indivíduos foram coletados por um museu sul-africano. Louis Albert Péringuey, taxonomista desse mesmo museu, examinou-os e chegou à conclusão de que eram de um novo gênero e espécie, mas não se sabe até hoje em que Ordem Louis os teria colocado, já que nunca foram formalmente classificados. Como consequência, foram engavetados juntos a outros louva-a-deus não identificados durante mais de 100 anos. No entanto, uma pesquisa recente redescobriu esses espécimes e os classificou corretamente após coletarem indivíduos vivos e chegarem a conclusão de que pertenciam a uma nova Ordem: Mantophasmatodea. Essa Ordem foi oficialmente descrita em 2002.
Morfologia editar
.jpg)
Mantophasmatodea podem ser facilmente confundidos com juvenis de louva-a-deus (ordem Mantodea), bicho-pau (ordem Phasmatodea), ou gafanhotos (ordem Orthoptera). Os adultos têm por volta de 2 a 3 centímetros de comprimento e são ápteros, ou seja, não possuem asas.
Superficialmente, eles se parecem com uma ninfa de louva-a-deus (que não tem asas nesse estágio), exceto pelo fato de não possuírem pernas raptoriais bem desenvolvidas e apresentarem antenas mais longas, com uma dobra terminal. A cabeça dirigida para baixo e o hábito alimentar os distinguem dos bichos-pau.
Mantophasmatodea não têm pernas traseiras saltatórias como a maioria dos gafanhotos e grilos, mas são tecnicamente separados de Orthoptera com base nos caracteres presentes no tórax e ovipositor. Sua notável semelhança com estágios juvenis de outras ordens de insetos é provavelmente o motivo pelo qual essa nova ordem foi negligenciada por muitos anos. Os espécimes vivos, no entanto, são facilmente reconhecidos pelo seu hábito de manter o último segmento do tarso (segmento mais distal da perna, ligado à tíbia, articulado, constituído por artículos denominados tarsômeros. Ver figura 3) e o arólio (estrutura eversível que auxilia a aderência do inseto ao substrato. Ver figura 4) no ar, sem entrar em contato com o substrato.
{kind=link}

Possuem três pares de pernas, corpo e extremidades espinhosas (Ver figura 2). A cabeça é hipognata (peças bucais estão voltadas para baixo), triangular em aspecto anterior e com bordas arredondadas, e o aparelho bucal é do tipo mastigador. Possuem mandíbulas com dois ou três dentículos (dentes pequenos) em área molar e olhos proeminentes em forma de rim, além de duas antenas longas, filiformes, multissegmentadas e ocelos (estrutura localizada na face dorsal da cabeça, anteriormente, responsável pela captação de luz) ausentes.
O tórax dos insetos é dividido em três partes: protórax (1º segmento do tórax, ligado à cabeça, onde está inserido o primeiro par de pernas), mesotórax (2º segmento do tórax, onde se encontra o segundo par de pernas e o primeiro par de asas, quando presentes) e o metatórax (3º segmento do tórax, ligado ao abdome, onde estão situados o terceiro par de patas e o segundo par de asas, quando presentes). Em Mantophasmatodea, cada parte dessas é sobreposta à seguinte (Ver figura 2).
O protórax e o mesotórax são hexagonais na fase adulta. O protórax possui margens anteriores com dois grandes espinhos sub-lateralmente e dois menores submedialmente. As margens laterais possuem dois grandes espinhos medialmente e dois pequenos posteriormente. A margem posterior é sobreposta à margem anterior do mesotórax, com seis grandes espinhos. O mesotórax é tão longo quanto o protórax, com uma linha mais escura medialmente. Sua margem anterior fica oculta abaixo da margem posterior do protórax. A margem lateral possui um espinho anteriormente e um posteriormente. Há dois grandes espinhos adjacentes às margens medialmente. A margem posterior possui seis grandes espinhos. O metatórax é consideravelmente mais curto do que o mesotórax. A margem anterior não contém espinhos, e se encontra escondida abaixo da margem posterior do mesotórax. A margem lateral possui um espinho pequeno. As pernas são espinhosas, com garras dirigidas para fora.

As asas são totalmente ausentes e as coxas, alongadas. Tarso com 5 tarsômeros (Ver figura 3), sendo 4 basais com euplantulae (Ver figura 4), que são como “almofadas de calcanhar” ou "almofadas de fricção" especializadas que produzem tração quando pressionadas contra o substrato. Possuem um arólio (estruturas adesivas que aderem ao substrato quando ativadas por forças de tração, em forma de almofada, entre as garras ou unhas de alguns insetos. Ver figura 4) pré-tarsal muito grande, com fileiras de cerdas longas.
Os segmentos abdominais vão aumentando do I ao VI e diminuindo do VII ao VIII, em comprimento e largura. O segmento I possui uma fileira de quatro grandes espinhos posteriormente. Dos segmentos II ao VI há quatro grandes espinhos posteriores e um grande espinho sub-medial, que vão diminuindo em tamanho nos seguintes segmentos.[1]
As ninfas desses insetos são menores, mas as proporções do corpo e extremidades não mostram notável diferenças para os adultos. Os espinhos nas extremidades e no corpo não são produzidos nos estágios iniciais de ninfas e aumentam de tamanho a cada muda. No estágio anterior à maturidade, os espinhos são quase produzidos como nos adultos. O tórax é mais trapezoidal entre as ninfas jovens e as quase adultas; a forma hexagonal torna-se distinta nas últimas quatro mudas. Nos machos, a furca do cerco (apêndice unitário ou em par, localizado no final do abdome, normalmente com função sensorial) torna-se visível nas três etapas anteriores à maturidade.
As diferenças mais óbvias entre os sexos estão na estrutura da genitália externa. A fêmea tem um ovipositor complicado, enquanto o macho tem estruturas fálicas internas e cercos bem desenvolvidos, que são usados como cláspers (estrutura em insetos machos usada para segurar a fêmea durante a cópula). O abdômen feminino é mais largo no meio (onde os ovários estão situados) em contraste com o do macho, que é mais largo perto da ponta. Os machos são consideravelmente menores e sempre mais delgados que as fêmeas.
{kind=link}

Ambos os sexos possuem um arólio[2] muito grande no pré-tarso de todas as pernas, que se encontram, junto ao 5º tarsômero, em posição ereta (virados para cima e para fora do substrato); esta posição altamente característica é bastante singular entre os insetos. Estudos demonstraram que os arólios raramente são usados para caminhar e não são necessários para produzir, detectar ou responder a sinais de comunicação vibracional. Observa-se o uso do arólio quando é necessária força de adesão adicional, ou seja, para aderir a superfícies lisas, estabilizar sua posição após o pouso ou no manuseio de grandes presas. A adesão eficaz é provida por secreções produzidas em glândulas epiteliais existentes nessas estruturas[3]. A produção de secreção é tipicamente consumidora de energia, o que pode ser uma razão para manter o arólio em uma posição ascendente quando não estiver em uso.
{kind=link}
Várias espécies exibem polimorfismo cromático com espécimes na mesma população, variando dramaticamente na cor. Por exemplo, a população de Austrophasma caledonensis (África do Sul) contém espécimes que variam de marrom a verde e branco a verde, e as populações de Austrophasma gansbaaiensis e Lobophasma redelinghuysensis podem ter morfos verdes e marrons. Há excelente correspondência de substrato, já que as espécies com morfos verdes ocupam a vegetação mais esverdeada como pradarias, e as formas cinzentas ou pardas ocorrem em áreas mais áridas. Tanto machos como fêmeas exibem todos os morfos de cor. A camuflagem é importante para esses pequenos predadores, que são vulneráveis ao ataque de várias aves, lagartos e outros insetos que ocorrem no mesmo microhabitat.
Ecologia editar
São insetos carnívoros tanto com hábito noturno como diurno, dependendo da espécie. Espécies sul-africanas são noturnas, enquanto as da Namíbia são diurnas.
{kind=link}
Esse grupo está presente em micro habitats com grande densidade de artrópodes. Enquanto não estão no seu período ativo (de procura de alimentos) ficam escondidos em rochas e vegetações rasteiras (onde se camuflam muito bem para impedir que sejam predados), acenando suas antenas continuamente de maneira similar a baratas, ou seja, ambas antenas se movem alternadamente para cima e para baixo. Já a cintilação da antena foi observada em contato com um parceiro sexual em potencial ou com uma presa. Esse comportamento indica uso intensivo de sinais químicos.
Hábito Alimentar editar
Os chamados gladiadores caçam presas de diversos tipos, que podem variar desde moscas, baratas, grilos, pequenas mariposas, aranhas até traças e animais tão grandes quanto eles. Podem até mesmo praticar canibalismo. Eles utilizam o primeiro par de pernas, espessas e com espinhos, para lutar com as suas presas e capturá-las. Quando a sua presa é maior, pode segurá-la ao mesmo tempo com o seu segundo par de pernas, também portador de espinhos (gafanhotos carnívoros utilizam uma técnica de caça com os primeiros dois pares de pernas de maneira parecida). Quando caçam moscas, atacam primeiramente seus pescoços com fortes mordidas e devoram suas cabeças antes de qualquer outra parte do corpo. Eles devoram suas presas por inteiro, evitando apenas as asas e os apêndices.
Comportamento Sexual editar
Todas as espécies de Mantophasmatodea exibem forte dimorfismo sexual na estrutura genital externa[4], e os machos são, em média, menores e mais delgados em comparação às fêmeas. O sistema reprodutor masculino inclui testículos, ducto deferente, vesículas seminais e glândulas acessórias[5]. Dependendo da espécie, fêmeas grávidas têm de 8 a 15 ovos por ovário, que são rodeados por uma fina camada de tecido e tipicamente ovipositados em um único envoltório. O peso da fêmea aumenta notavelmente durante a maturação dos ovos.
As fêmeas ovipositam e produzem envoltórios formados por areia e um líquido secretado para proteger os ovos. Esses envoltórios endurecem devido à areia incorporada a eles e são colocados superficialmente (0,5 a 2 cm de profundidade) em solo arenoso. Os ovos exibem um arranjo vertical regular semelhante ao de garrafas em uma caixa de cerveja. Não é comum que espécies de Mantophasmatodea depositem ovos estéreis, diferentemente do que ocorre nas espécies de louva-a-deus[6].
{kind=link}
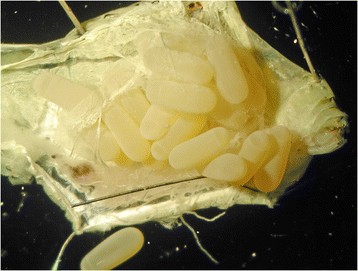{kind=link}
{kind=link}
Além da influência direta das chuvas no desenvolvimento dos ovos, não há informações sobre quais fatores adicionais induzem o fim da dormência após o período seco.
Comportamento de Percussão editar
Ambos os sexos usam chamadas vibracionais únicas, que são produzidas tocando-se o abdômen no substrato para localização e reconhecimento de posição.[7] A placa subgenital masculina tem um processo semicircular que temporariamente entra em contato com a superfície durante a percussão. As fêmeas não desenvolvem este processo abdominal e batem com a parte média do ventre. As chamadas femininas são pulsos repetidos únicos[8], enquanto as chamadas masculinas são pulsações mais complexas que provavelmente contêm codificação para reconhecimento de espécies.
Cortejo e Comportamento de Acasalamento editar
Se mantido em gaiolas espaçosas, o acasalamento em espécies de Austrophasmatidae começa principalmente à tarde ou à noite. Em contraste, o acasalamento de vários outros espécimes foi observado a qualquer hora do dia.
Eberhard e Picker [7] hipotetizaram que os machos eram atraídos quimicamente através de feromônios femininos por um longo alcance e através de comunicação vibracional contínua. No entanto, mesmo que a comunicação vibracional faça parte do comportamento sexual, não é um comportamento obrigatório para localizar um parceiro sexual em experimentos observados em laboratório. Parceiros sexuais pareciam notar a presença um do outro a aproximadamente 8 a 10 cm de distância. À medida que se aproximavam, ocorria uma rápida vibração de suas antenas, que indica comunicação sexual mediada por odor. Na maioria dos casos, as antenas não contatavam diretamente o parceiro[9], mas ocasionalmente, havia contato.
Foi frequentemente observado os machos movendo seu abdômen para cima e para baixo, enquanto as fêmeas se aproximavam deles ou batiam continuamente seus abdomens na superfície.
Em um caso observado, uma fêmea do Hemilobophasma montaguense caminhava de um lado para o outro na frente de um macho. De repente, a fêmea parou e moveu seu abdome para cima e, imediatamente, o macho se aproximou e montou. Os machos montam rapidamente. Durante a cópula, eles seguram as fêmeas com suas pernas dianteiras e médias. Logo após a montagem, eles retêm seu arólio para cima.[7]. Para a cópula, estendem seu abdômen para o lado direito das fêmeas, formando um “s”, enquanto elas projetam seu abdome para cima. Os cercos dos machos são usados como cláspers para aproximar a genitália de ambos. A grande genitália masculina evagina e se insere na vagina. Durante o acasalamento prolongado, os machos permanecem montados, independentemente do comportamento da fêmea. Durante a cópula, foi observado fêmeas caminhando, perseguindo presas e excretando[7] Mesmo que uma fêmea caísse, o macho permanecia sentado em seu abdome. Ocasionalmente, foram encontrados machos montados em fêmeas mortas.
{kind=link}
Os machos geralmente parecem estar em risco de ataque pela fêmea, como indicado pela sua rápida montagem. Zompro et al[1] relataram que os machos de Mantophasma kudubergense recuaram pulando para longe se as fêmeas parecessem agitadas após a montagem.
Os elementos de comportamento sexual, tais como percussão, antenas piscando por sinais químicos e contato com antenas, podem ser interpretados como comportamento de reconhecimento de espécie[7] ou comportamento de corte para reduzir ataques femininos; os elementos podem estar envolvidos em ambos os comportamentos.
Foi observada uma substância gelatinosa semilíquida entre as estruturas genitais externas de machos e fêmeas durante a última fase da cópula, que foi assumida como uma substância liberada pelo macho e que era um tipo de plugue de acasalamento, e não um espermatóforo.
Custos de Acasalamento editar
Em cativeiro, as fêmeas em cópula eram regularmente observadas caçando e alimentando-se de insetos. Portanto, os custos do acasalamento podem ser baixos para as fêmeas. Em contraste, os machos não comem durante a cópula, perdendo assim uma quantidade significativa de peso. Assume-se que um curto período de inanição é um fenômeno natural em Mantophasmatodea. Além da fome e desidratação, a transferência de quantidades substanciais de proteínas e líquidos dos machos para as fêmeas também deve ser considerada.
Desenvolvimento editar
A ordem Mantophasmatodea está inserida na superordem Polyneoptera e portanto são hemimetábolos, modo de desenvolvimento que passa por três fases distintas: o ovo, a ninfa e o estágio adulto ou imago. Os Mantophasmatodea se reproduzem uma vez por ano.
As informações a respeito da cópula desse animal ainda são muito rasas e não foram descritas com precisão. Após a fecundação, as fêmeas depositam os ovos rapidamente caso haja solo árido disponível, aproximadamente 30 capsulas com ovos são depositadas por fêmea, cada uma delas com um grande número de ovos fecundados dentro, aproximadamente de trinta a cinquenta ovos por vagem[10].
Os ovos entram em fase de eclosão apenas na estação chuvosa. Diversos testes foram realizados expondo os ovos à diversas variações climáticas e nenhuma delas induziu a eclosão, exceto as que simulavam a estação chuvosa[11]. Devido à sua alta resistência, os ovos podem apresentar estado de dormência e sobrevivem por pelo menos dois anos após a oviposição, algo em comum com uma espécie da ordem Orthoptera[12]. Aproximadamente depois de um ano os ovos passam pela estação chuvosa e as ninfas eclodem, atingindo a idade adulta depois de três a cinco meses.[9] Como após as ecdises a exúvia é consumida pelo animal, ainda não se sabe por quantas mudas o animal passa até atingir o estado adulto, ou seja, até atingir o estado de desenvolvimento pleno dos órgãos reprodutivos.
Os adultos vivem em média cinco meses e portanto passam por um período de reprodução. As fêmeas costumam viver por mais tempo e podem realizar duas reproduções em um mesmo ano se condições de muita chuva se manterem constantes.Ainda não há informações disponíveis sobre fatores de mortalidade, como predação e parasitismo,pois devido a recente descoberta do grupo, os estudos em campo ainda não foram realizados. Supõe-se que a camuflagem seja importante para esses pequenos predadores, que provavelmente são vulneráveis ao ataque de pássaros, lagartos e insetos carnívoros presentes no mesmo hábitat. O comportamento de fuga, isto é, saltar de um arbusto, não é uma característica dos insetos gladiadores.
Distribuição editar
São espécies endêmicas. Representantes do grupo foram encontrados na Namíbia (família: Mantophasmatidae; Gênero: Tyrannophasma; Gênero: Praedatophasma), Tanzânia (Família: Tanzaniophasmatidae) e nas regiões norte e oeste da Cidade do Cabo, na África do sul (Família: Austrophasmatidae).
Também foi encontrado um gênero já extinto chamado Rhaptophasma, em âmbar. Os fósseis são de aproximadamente 45 milhões de anos (época do eoceno) e foram localizados na região do Mar Báltico e Península Escandinávia, na Europa.
Taxonomia e Filogenia editar
| Classificação Mantophasmatodea dentro dos Polyneoptera | |
|
| |
| Cladograma baseado em Gullan & Cranston (2008). |
A recente descoberta dessa ordem[1] e a quantidade baixa de espécies conhecidas causa problemas na classificação baseada em características morfológicas. Entretanto baseado em estudos moleculares recentes foi possível classificá-lo como grupo irmão de Gryllobattodea[13] e os dois grupos juntos formam a ordem maior Notoptera [14]. Os Mantophasmatodea estão contidos na subclasse Polyneoptera, com desenvolvimento hemimetábolo, não sofrendo grandes mudanças na forma do corpo e comportamento ao longo de sua vida. A subclasse Polyneoptera contém ordens com diversas perdas secundárias de aparelhos morfológicos especializados como as asas, condição que possibilita o desenvolvimento de diversas hipóteses filogenéticas. Seu nome é uma junção dos nomes Mantodea e Phasmatodea, grupos presentes na mesma subclasse, devido sua similaridade morfológica.
Com o baixo número de espécies catalogadas os insetos gladiadores possuem poucas famílias e gêneros, dificultando a classificação dentro da ordem. Ao todo atualmente foram descobertas quatorze espécies desse animal, sendo colocadas em onze gêneros e quatro famílias[15]. Estudos mais atuais revelam que há provavelmente muitas espécies não descobertas e com isso novas classificações serão criadas.
| Família | Gêneros | Espécies |
|---|---|---|
| Tanzaniophasmatidae | Tanzaniophasma | Tanzaniophasma subsolana |
| Mantophasmatidae | Mantophasma | Mantophasma zephyra |
| Sclerophasma | Sclerophasma paresisensis | |
| Austrophasmatidae | Austrophasma | Austrophasma caledonensis |
| Austrophasma gansbaaiensis | ||
| Austrophasma rawsonvillensis | ||
| Lobophasma | Lobophasma redelinghuysensis | |
| Hemilobophasma | Hemilobophasma montaguensis | |
| Karoophasma | Karoophasma biedouwensis | |
| Karoophasma botterkloofensis | ||
| Namaquaphasma | Namaquaphasma ookiepensis | |
| Inquirendo | Rhaptophasma | Rhaptophasma kerneggeri |
| Praedatophasma | Praedatophasma maraisi | |
| Tyrannophasma | Tyrannophasma gladiator |
Referências editar
- ↑ a b c Klass, Klaus-D.; Zompro, Oliver; Kristensen, Niels P.; Adis, Joachim (24 de maio de 2002). «Mantophasmatodea: A New Insect Order with Extant Members in the Afrotropics». Science (em inglês). 296 (5572): 1456–1459. ISSN 0036-8075. PMID 11964441. doi:10.1126/science.1069397
- ↑ «Entomologia - Glossário UFRGS». www.ufrgs.br. UFRGS. Setembro de 2007. Consultado em 7 de junho de 2018
- ↑ Eberhard, Monika J.B.; Pass, Günther; Picker, Mike D.; Beutel, Rolf; Predel, Reinhard; Gorb, Stanislav N. (outubro de 2009). «Structure and function of the arolium of Mantophasmatodea (Insecta)». Journal of Morphology (em inglês). 270 (10): 1247–1261. ISSN 0362-2525. doi:10.1002/jmor.10754. Consultado em 10 de junho de 2018
- ↑ KLASS, KD (12 de setembro de 2003). «The taxonomy, genitalic morphology, and phylogentic relationships of southern African Mantophasmatodea (Insecta)» (PDF). Entomologische Abhandlungen. Consultado em 12 de junho de 2018
- ↑ Dallai, Romano; Lupetti, Pietro; Afzelius, Bj�rn A.; Frati, Francesco (1 de novembro de 2003). «Sperm structure of Mecoptera and Siphonaptera (Insecta) and the phylogenetic position of Boreus hyemalis». Zoomorphology (em inglês). 122 (4): 211–220. ISSN 0720-213X. doi:10.1007/s00435-003-0087-y replacement character character in
|primeiro3=at position 3 (ajuda) - ↑ CUKIER, MARTA; GUERRERO, GRACIELA ALICIA; MAGGESE, MARÍA CRISTINA (dezembro de 1979). «PARTHENOGENESIS IN COPTOPTERYX VIRIDIS, GIGLIO TOS (1915) (DYCTIOPTERA, MANTIDAE)». The Biological Bulletin (em inglês). 157 (3): 445–452. ISSN 0006-3185. doi:10.2307/1541029
- ↑ a b c d e Eberhard, Monika J. B.; Picker, Mike D. (15 de abril de 2008). «Vibrational Communication in Two Sympatric Species of Mantophasmatodea (Heelwalkers)». Journal of Insect Behavior (em inglês). 21 (4): 240–257. ISSN 0892-7553. doi:10.1007/s10905-008-9123-6
- ↑ Eberhard, Monika J. B.; Eberhard, Stefan H. (12 de agosto de 2012). «Evolution and Diversity of Vibrational Signals in Mantophasmatodea (Insecta)». Journal of Insect Behavior (em inglês). 26 (3): 352–370. ISSN 0892-7553. doi:10.1007/s10905-012-9352-6
- ↑ a b Zompro, Oliver; Adis, Joachim; E. Bragg, Philip; Naskrecki, Piotr; Meakin, Kathy; Wittneben, Martin; Saxe, Victoria (1 de janeiro de 2003). «A new genus and species of Mantophasmatidae (Insecta: Mantophasmatodea) from the Brandberg Massif, Namibia, with notes on behaviour». Cimbebasia, v.19, 13-24 (2003). 19
- ↑ Uchifune T: Collection and rearing of a South African heel-walker, Karoophasma biedouwensis Klass et al. (Insecta: Mantophasmatodea). Science Report of the Yokosuka City Museum. 2008, 55: 23-28.
- ↑ Roth, Steffen; Molina, Jorge; Predel, Reinhard (10 de outubro de 2014). «Biodiversity, ecology, and behavior of the recently discovered insect order Mantophasmatodea». Frontiers in Zoology. 11 (1). 70 páginas. ISSN 1742-9994. doi:10.1186/s12983-014-0070-0
- ↑ Power, J.H. (1 de novembro de 1958). «On The Biology of Acanthoplus bechuanus Per. (Orthoptera: Tettigoniidae)». Journal of the Entomological Society of Southern Africa (em inglês). 21 (2). ISSN 0013-8789
- ↑ «Evidence fron Mitochondrial Genomics on Interordinal Relationships in Insects»
- ↑ «Rock Crawlers in Baltic Amber (Notoptera: Mantophasmatodea)» (PDF) line feed character character in
|titulo=at position 30 (ajuda) - ↑ «Mantophasmatodea (heelwalkers, mantos)». www.biodiversityexplorer.org. Consultado em 28 de maio de 2018