Plexo coroide
Plexo coroide (do latim plexu, enlaçamento, e do grego choroeidés, membrana delicada), é uma estrutura encontrada nos ventrículos do sistema nervoso[1] onde é produzido a maior parte do líquido cefalorraquidiano (LCR).[2] O LCR é produzido e secretado por diferentes regiões do plexo coróide.[3] O plexo coróide consiste em células ependimárias modificadas ao redor de um núcleo de capilares e tecido conjuntivo frouxo . Num adulto, ocorre produção de cerca de 500 ml/dia e usado para drenar certas substâncias (como drogas e toxinas) para fora do cérebro. Todo o volume de LCR é trocado em torno de três a quatro vezes por dia.[4]

É resultado de invaginação do teto do epêndima para a cavidade ventricular. Possui epitélio cuboide ou colunar simples e ocupa cerca de 40cm² de área. Não possui a barreira hematoencefálica típica do encéfalo, o que os tornam vulneráveis a doenças sistêmicas.[4]
Lesões expansivas do plexo coróide estão associadas a casos de hidrocefalia, com maior incidência em crianças.[4]
Estrutura
editarLocalização
editar
Existe um plexo coróide em cada um dos quatro ventrículos. Nos ventrículos laterais, é encontrado no corpo e continua em quantidade aumentada no átrio. Não há plexo coróide no corno anterior No terceiro ventrículo existe uma pequena porção no teto que é contínua com a do corpo, via forames interventriculares, os canais que conectam os ventrículos laterais com o terceiro ventrículo. Há um plexo coróide em parte do teto do quarto ventrículo .
Microanatomia
editarO plexo coróide consiste em uma camada de células epiteliais cuboidais circundando um núcleo de capilares e tecido conjuntivo frouxo.[3] O epitélio do plexo coróide é contínuo com a camada de células ependimárias (camada ventricular) que reveste o sistema ventricular. As células ependimárias progenitoras são monociliadas, mas se diferenciam em células ependimárias multiciliadas.[5][6] Ao contrário do epêndima, a camada epitelial do plexo coróide tem junções de oclusão[7] entre as células no lado voltado para o ventrículo (superfície apical). Essas junções estreitas impedem que a maioria das substâncias atravesse a camada celular para o líquido cefalorraquidiano (LCR); assim, o plexo coróide atua como uma barreira sangue-LCR. O plexo coróide se dobra em muitas vilosidades ao redor de cada capilar, criando processos semelhantes a folhas que se projetam nos ventrículos. As vilosidades, junto com uma borda em escova de microvilosidades, aumentam muito a área de superfície do plexo coróide. CSF é formado como plasma é filtrado do sangue através das células epiteliais. As células epiteliais do plexo coróide transportam ativamente íons de sódio para os ventrículos e a água segue o gradiente osmótico resultante.[8]
O plexo coróide consiste em muitos capilares, separados dos ventrículos por células epiteliais coróides. O líquido filtra essas células do sangue para se tornar líquido cefalorraquidiano. Há também muito transporte ativo de substâncias para dentro e para fora do CSF à medida que ele é feito.

O plexo coróide regula a produção e composição do líquido cefalorraquidiano (LCR), que fornece uma flutuabilidade protetora para o cérebro.[2][9] O LCR atua como um meio para o sistema de filtração glinfática que facilita a remoção de resíduos metabólicos do cérebro e a troca de biomoléculas e xenobióticos para dentro e para fora do cérebro.[9][10] Desta forma, o plexo coróide tem um papel muito importante em ajudar a manter o delicado ambiente extracelular exigido pelo cérebro para funcionar de forma otimizada.
O plexo coróide também é uma importante fonte de secreção de transferrina, que desempenha um papel na homeostase do ferro no cérebro.[11][12]
Barreira sangue-líquido cefalorraquidiano
editarA barreira hematoencefálica (BCSFB) é uma barreira hematoencefálica composta por um par de membranas que separam o sangue do LCR em nível capilar e o LCR do tecido cerebral.[13] O limite sangue-LCR no plexo coróide é uma membrana composta de células epiteliais conectadas por junções de oclusão.[13] Existe uma barreira LCR-cérebro ao nível da pia-máter, mas apenas no embrião.[14]
Semelhante à barreira hematoencefálica, a barreira sangue-líquido cefalorraquidiano funciona para prevenir a passagem da maioria das substâncias do sangue para o cérebro, enquanto permite seletivamente a passagem de substâncias específicas para o cérebro e facilita a remoção de metabólitos cerebrais e produtos metabólicos para o sangue.[13][15] Apesar da função semelhante entre o BBB e o BCSFB, cada um facilita o transporte de diferentes substâncias para o cérebro devido às características estruturais distintas de cada um dos dois sistemas de barreira.[13] Para várias substâncias, o BCSFB é o principal local de entrada no tecido cerebral.[13]
A barreira sangue-líquido cefalorraquidiano também modula a entrada de leucócitos do sangue para o sistema nervoso central. As células do plexo coróide secretam citocinas que recrutam macrófagos derivados de monócitos, entre outras células, para o cérebro. Esse tráfego celular tem implicações tanto na homeostase normal do cérebro quanto nos processos neuroinflamatórios .[16]
Significado clínico
editarCistos do plexo coróide
editarDurante o desenvolvimento fetal, alguns cistos do plexo coróide podem se formar. Esses cistos cheios de líquido podem ser detectados por um ultrassom detalhado do segundo trimestre. O achado é relativamente comum, com uma prevalência de ~ 1%. Os cistos do plexo coróide são geralmente um achado isolado.[17] Os cistos geralmente desaparecem mais tarde durante a gravidez e geralmente são inofensivos. Eles não têm efeito sobre o desenvolvimento do bebê e da primeira infância.[18]
Cistos conferem um risco de 1% de aneuploidia fetal.[19] O risco de aneuploidia aumenta para 10,5-12% se outros fatores de risco ou achados ultrassonográficos forem observados. Tamanho, localização, desaparecimento ou progressão, e se os cistos são encontrados em ambos os lados ou não, não afetam o risco de aneuploidia. 44-50% dos casos de síndrome de Edwards (trissomia do cromossomo 18) se apresentarão com cistos do plexo coróide, assim como 1,4% dos casos de síndrome de Down (trissomia do cromossomo 21). ~ 75% dos cariótipos anormais associados a cistos do plexo coróide são trissomia 18, enquanto o restante é trissomia 21.[17]
Etimologia
editarPlexo coróide é traduzido do latim plexo chorioides,[20] que reflete o grego antigo χοριοειδές πλέγμα.[21] A palavra córion foi usada por Galeno para se referir à membrana externa que envolve o feto. Ambos os significados da palavra plexo são dados como pregas ou tranças.[21] Tanto o uso de coróide como corioide são aceitos. A Nomina Anatomica (agora Terminologia Anatomica) refletia esse duplo uso.
Imagens adicionais
editar-
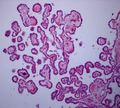 Histologia do plexo coróide 40x
Histologia do plexo coróide 40x -
 Plexo coróide
Plexo coróide -
 Plexo coróide
Plexo coróide -
 Plexo coróide
Plexo coróide




Referências
- ↑ Sadler, T. (2010). Langman's medical embryology 11th ed. Philadelphia: Lippincott William & Wilkins. ISBN 978-0-7817-9069-7
- ↑ a b Damkier, HH; Brown, PD; Praetorius, J (outubro de 2013). «Cerebrospinal fluid secretion by the choroid plexus.» (PDF). Physiological Reviews. 93: 1847–92. PMID 24137023. doi:10.1152/physrev.00004.2013
- ↑ a b Lun, MP; Monuki, ES; Lehtinen, MK (agosto de 2015). «Development and functions of the choroid plexus-cerebrospinal fluid system.». Nature Reviews. Neuroscience. 16: 445–57. PMC 4629451
 . PMID 26174708. doi:10.1038/nrn3921
. PMID 26174708. doi:10.1038/nrn3921
- ↑ a b c MELO, Ernesto Lima Araújo et al. Lesões expansivas do plexo coróide. Radiol Bras [online]. 2003, vol.36, n.6 [cited 2013-08-10], pp. 379-384 . Available from: <http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-39842003000600010&lng=en&nrm=iso>. ISSN 0100-3984. http://dx.doi.org/10.1590/S0100-39842003000600010.
- ↑ Delgehyr, N; Meunier, A; Faucourt, M; Bosch Grau, M; Strehl, L; Janke, C; Spassky, N (2015). «Ependymal cell differentiation, from monociliated to multiciliated cells.». Methods in Cell Biology. 127: 19–35. ISBN 9780128024515. PMID 25837384. doi:10.1016/bs.mcb.2015.01.004
- ↑ van Leeuwen LM, Evans RJ, Jim KK, Verboom T, Fang X, Bojarczuk A, Malicki J, Johnston SA, van der Sar AM (fevereiro de 2018). «A transgenic zebrafish model for the in vivo study of the blood and choroid plexus brain barriers using claudin 5». Biology Open. 7: bio030494. PMC 5861362 . PMID 29437557. doi:10.1242/bio.030494
- ↑ Hall, John (2011). Guyton and Hall textbook of medical physiology 12th ed. Philadelphia, Pa.: Saunders/Elsevier. ISBN 978-1-4160-4574-8
- ↑ Guyton, Arthur C.; Hall, John Edward (2005). Textbook of medical physiology 11th ed. Philadelphia: W.B. Saunders. pp. 764–7. ISBN 978-0-7216-0240-0
- ↑ a b Plog BA, Nedergaard M (janeiro de 2018). «The Glymphatic System in Central Nervous System Health and Disease: Past, Present, and Future». Annual Review of Pathology. 13: 379–394. PMC 5803388 . PMID 29195051. doi:10.1146/annurev-pathol-051217-111018
- ↑ Abbott NJ, Pizzo ME, Preston JE, Janigro D, Thorne RG (março de 2018). «The role of brain barriers in fluid movement in the CNS: is there a 'glymphatic' system?». Acta Neuropathologica. 135: 387–407. PMID 29428972. doi:10.1007/s00401-018-1812-4
- ↑ Moos, T (novembro de 2002). «Brain iron homeostasis». Danish Medical Bulletin. 49: 279–301. PMID 12553165
- ↑ Moos, T; Rosengren Nielsen, T; Skjørringe, T; Morgan, EH (dezembro de 2007). «Iron trafficking inside the brain». Journal of Neurochemistry. 103: 1730–40. PMID 17953660. doi:10.1111/j.1471-4159.2007.04976.x
- ↑ a b c d e «Blood–cerebrospinal fluid barrier». Basic Neurochemistry: Molecular, Cellular and Medical Aspects 6th ed. Philadelphia: Lippincott-Raven. 1999
- ↑ Saunders, Norman R.; Habgood, Mark D.; Møllgård, Kjeld; Dziegielewska, Katarzyna M. (10 de março de 2016). «The biological significance of brain barrier mechanisms: help or hindrance in drug delivery to the central nervous system?». F1000Research. 5. 313 páginas. ISSN 2046-1402. PMC 4786902 . PMID 26998242. doi:10.12688/f1000research.7378.1.
The embryonic CSF-brain barrier, shown in Figure 1(f). In the ventricular zone is a temporary barrier between the CSF and brain parenchyma. In early brain development, strap junctions are present between adjacent neuroepithelial cells; these form a physical barrier restricting the movement of larger molecules, such as proteins, but not smaller molecules. At later stages of development and in the adult brain, these strap junctions are no longer present when this interface becomes ependyma.
- ↑ Ueno M, Chiba Y, Murakami R, Matsumoto K, Kawauchi M, Fujihara R (abril de 2016). «Blood-brain barrier and blood-cerebrospinal fluid barrier in normal and pathological conditions». Brain Tumor Pathology. 33: 89–96. PMID 26920424. doi:10.1007/s10014-016-0255-7
- ↑ Schwartz M, Baruch K (janeiro de 2014). «The resolution of neuroinflammation in neurodegeneration: leukocyte recruitment via the choroid plexus». The EMBO Journal. 33: 7–22. PMC 3990679 . PMID 24357543. doi:10.1002/embj.201386609
- ↑ a b Drugan A, Johnson MP, Evans MI (janeiro de 2000). «Ultrasound screening for fetal chromosome anomalies». American Journal of Medical Genetics. 90: 98–107. PMID 10607945. doi:10.1002/(SICI)1096-8628(20000117)90:2<98::AID-AJMG2>3.0.CO;2-H
- ↑ Digiovanni LM, Quinlan MP, Verp MS (agosto de 1997). «Choroid plexus cysts: infant and early childhood developmental outcome». Obstetrics and Gynecology. 90: 191–4. PMID 9241291. doi:10.1016/S0029-7844(97)00251-2
- ↑ Peleg D, Yankowitz J (julho de 1998). «Choroid plexus cysts and aneuploidy». Journal of Medical Genetics. 35: 554–7. PMC 1051365 . PMID 9678699. doi:10.1136/jmg.35.7.554
- ↑ Suzuki, S., Katsumata, T., Ura, R. Fujita, T., Niizima, M. & Suzuki, H. (1936). Über die Nomina Anatomica Nova. Folia Anatomica Japonica, 14, 507-536.
- ↑ a b A Greek-English Lexicon. Oxford: Clarendon Press. 1940
- Este artigo incorpora texto em domínio público da 20ª edição de Gray's Anatomy (1918)