Potencial elétrico de membrana
Potencial elétrico de membrana (ou potencial transmembranar ou voltagem da membrana) é a diferença de potencial eléctrico (voltagem) entre os meios intra e extracelular. Ou seja, há uma diferença na energia necessária para que as cargas elétricas se movam do ambiente celular interno para o externo e vice-versa, desde que não haja aquisição de energia cinética ou produção de radiação. Os gradientes de concentração das cargas determinam diretamente essa necessidade de energia. Para o exterior da célula, os valores típicos de potencial, normalmente dados em unidades de milivolts e denotados como mV, variam de –80 mV a –40 mV.

Todas as células animais são envolvidas por uma membrana constituída por uma bicamada lipídica que contém proteínas incorporadas. A membrana celular serve tanto como isolante quanto como barreira de difusão para o movimento de íons. As bombas de íons, que são proteínas transmembranares, empurram os íons para que cruzem a membrana, através dos canais iônicos, assim modificando os gradientes de concentração. Bombas iónicas e canais iônicos são eletricamente equivalentes a um conjunto de pilhas e resistores inseridos na membrana e criam uma diferença de tensão entre os dois lados da membrana, regulando as concentrações de íons no citoplasma, separado pela bicapa lipídica da membrana plasmática.
Quase todas as membranas plasmáticas têm um potencial elétrico através delas, com o interior geralmente negativo em relação ao exterior.[1] O potencial de membrana tem duas funções básicas. Primeiro, permite que uma célula funcione como uma bateria, fornecendo energia para operar uma variedade de "dispositivos moleculares" embutidos na membrana.[2] Em segundo lugar, em células eletricamente excitáveis, como neurônios e células musculares, é usado para transmitir sinais entre diferentes partes de uma célula. Os sinais são gerados pela abertura ou fechamento de canais iônicos em um ponto da membrana, produzindo uma mudança local no potencial de membrana. Essa mudança no campo elétrico pode ser rapidamente percebida por canais iônicos adjacentes ou mais distantes na membrana. Esses canais iônicos podem então abrir ou fechar como resultado da mudança potencial, reproduzindo o sinal.
Em células não-excitáveis, e em células excitáveis em seus estados basais, o potencial de membrana é mantido em um valor relativamente estável, chamado potencial de repouso. Para os neurônios, o potencial de repouso é definido como variando de –80 a –70 milivolts. A abertura e o fechamento dos canais iônicos podem induzir um afastamento do potencial de repouso. Isso é chamado de despolarização se a voltagem interna se tornar menos negativa (digamos, de –70 mV a –60 mV), ou hiperpolarização se a voltagem interna se tornar mais negativa (digamos, de –70 mV a –80 mV). Em células excitáveis, uma despolarização suficientemente grande pode evocar um potencial de ação, no qual o potencial de membrana muda rápida e significativamente por um curto período de tempo (da ordem de 1 a 100 milissegundos), muitas vezes invertendo sua polaridade. Os potenciais de ação são gerados pela ativação de certos canais iônicos dependentes de voltagem.
Nos neurônios, os fatores que influenciam o potencial de membrana são diversos. Eles incluem vários tipos de canais iônicos, alguns dos quais são quimicamente controlados por neurotransmissores e alguns dos quais são dependentes de voltagem. Como os canais iônicos dependentes de voltagem são controlados pelo potencial de membrana, enquanto o próprio potencial de membrana é influenciado por esses mesmos canais iônicos, surgem loops de feedback que permitem dinâmicas temporais complexas, incluindo oscilações e eventos regenerativos, como potenciais de ação.
Base física
editarO potencial de membrana em uma célula deriva de dois fatores: força elétrica e difusão. A força elétrica surge da atração mútua entre partículas com cargas elétricas opostas (positivas e negativas) e da repulsão mútua entre partículas com o mesmo tipo de carga (ambas positivas ou ambas negativas). A difusão surge da tendência estatística das partículas de redistribuir de regiões onde estão altamente concentradas para regiões onde a concentração é baixa.
Voltagem
editar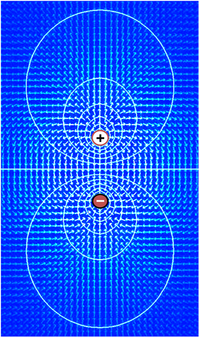
A tensão elétrica, que é sinônimo de diferença de potencial elétrico, é a capacidade de conduzir uma corrente elétrica através de uma resistência. De fato, a definição mais simples de tensão é dada pela lei de Ohm: V=IR, onde V é tensão, I é corrente e R é resistência. Se uma fonte de tensão, como uma bateria, for colocada em um circuito elétrico, quanto maior a tensão da fonte, maior a quantidade de corrente que ela conduzirá através da resistência disponível. O significado funcional da tensão reside apenas nas diferenças de potencial entre dois pontos em um circuito. A ideia de uma tensão em um único ponto não tem sentido. É convencional em eletrônica atribuir uma tensão de zero a algum elemento do circuito escolhido arbitrariamente e, em seguida, atribuir tensões para outros elementos medidos em relação a esse ponto zero. Não há significância em qual elemento é escolhido como o ponto zero - a função de um circuito depende apenas das diferenças e não das tensões em si. No entanto, na maioria dos casos e por convenção, o nível zero é mais frequentemente atribuído à parte de um circuito que está em contato com o solo.
O mesmo princípio se aplica à voltagem na biologia celular. Em tecido eletricamente ativo, a diferença de potencial entre quaisquer dois pontos pode ser medida inserindo um eletrodo em cada ponto, por exemplo, um dentro e um fora da célula, e conectando ambos os eletrodos aos terminais do que é essencialmente um voltímetro especializado. Por convenção, o valor de potencial zero é atribuído ao exterior da célula e o sinal da diferença de potencial entre o exterior e o interior é determinado pelo potencial do interior em relação ao zero exterior.
Em termos matemáticos, a definição de tensão começa com o conceito de um campo elétrico E, um campo vetorial que atribui uma magnitude e direção a cada ponto no espaço. Em muitas situações, o campo elétrico é um campo conservativo, o que significa que pode ser expresso como o gradiente de uma função escalar V, isto é, E = –∇V. Este campo escalar V é referido como a distribuição de tensão. Observe que a definição permite uma constante arbitrária de integração - é por isso que os valores absolutos de tensão não são significativos. Em geral, os campos elétricos podem ser tratados como conservadores apenas se os campos magnéticos não os influenciarem significativamente, mas essa condição geralmente se aplica bem ao tecido biológico.
Como o campo elétrico é o gradiente da distribuição de tensão, mudanças rápidas de tensão dentro de uma pequena região implicam em um forte campo elétrico; pelo contrário, se a tensão permanece aproximadamente a mesma em uma grande região, os campos elétricos nessa região devem ser fracos. Um forte campo elétrico, equivalente a um forte gradiente de voltagem, implica que uma forte força é exercida sobre quaisquer partículas carregadas que estejam dentro da região.
Íons e as forças que conduzem seu movimento
editarSinais elétricos dentro de organismos biológicos são, em geral, conduzidos por íons. Os cátions mais importantes para o potencial de ação são sódio (Na+) e potássio (K+).[3] Ambos são cátions monovalentes que carregam uma única carga positiva. Os potenciais de ação podem também envolver o cálcio (Ca2+),[3] que, por sua vez, é um cátion divalente que carrega uma carga positiva dupla. O ânion cloreto (Cl−) desempenha um papel importante nos potenciais de ação de algumas algas,[4] mas desempenha um papel negligenciável na maioria dos animais.
Os íons atravessam a membrana celular sob duas influências: difusão e campos elétricos. Um exemplo simples em que duas soluções – A e B – são separadas por uma barreira porosa ilustra que a difusão garantirá que elas eventualmente se misturarão em soluções iguais. Essa mistura ocorre por causa da diferença em suas concentrações. A região com alta concentração se difundirá em direção à região com baixa concentração. Para estender o exemplo, seja a solução A com 30 íons sódio e 30 íons cloreto. Além disso, deixe a solução B ter apenas 20 íons de sódio e 20 íons de cloreto. Assumindo que a barreira permite que ambos os tipos de íons passem por ela, então um estado estacionário será alcançado em que ambas as soluções terão 25 íons de sódio e 25 íons de cloreto. Se, no entanto, a barreira porosa for seletiva para os íons que passam, então a difusão por si só não determinará a solução resultante. Voltando ao exemplo anterior, vamos agora construir uma barreira permeável apenas aos íons sódio. Agora, apenas o sódio é permitido atravessar a barreira de sua concentração mais alta na solução A para a concentração mais baixa na solução B. Isso resultará em um acúmulo maior de íons sódio do que íons cloreto na solução B e um número menor de íons sódio do que íons cloreto na solução A .
Isso significa que há uma carga líquida positiva na solução B da concentração mais alta de íons sódio carregados positivamente do que íons cloreto carregados negativamente. Da mesma forma, há uma carga líquida negativa na solução A da maior concentração de íons cloreto negativos do que íons sódio positivos. Como cargas opostas se atraem e cargas semelhantes se repelem, os íons agora também são influenciados por campos elétricos, bem como por forças de difusão. Portanto, os íons de sódio positivos terão menos probabilidade de viajar para a solução B, agora mais positiva, e permanecer na solução A, agora mais negativa. O ponto em que as forças dos campos elétricos neutralizam completamente a força devido à difusão é chamado de potencial de equilíbrio. Neste ponto, o fluxo líquido do íon específico (neste caso sódio) é zero.
Membranas plasmáticas
editar
Cada célula é envolvida por uma membrana plasmática, que tem a estrutura de uma bicamada lipídica com muitos tipos de moléculas incorporadas nela. Por ser constituída de moléculas lipídicas, a membrana plasmática possui intrinsecamente uma alta resistividade elétrica, ou seja, uma baixa permeabilidade intrínseca a íons. No entanto, algumas das moléculas incorporadas na membrana são capazes de transportar ativamente íons de um lado da membrana para o outro ou fornecer canais através dos quais eles podem se mover.[5]
Na terminologia elétrica, a membrana plasmática funciona como um resistor e um capacitor combinados. A resistência surge do fato de que a membrana impede o movimento de cargas através dela. A capacitância surge do fato de que a bicamada lipídica é tão fina que um acúmulo de partículas carregadas de um lado dá origem a uma força elétrica que puxa partículas de carga oposta para o outro lado. A capacitância da membrana é relativamente não afetada pelas moléculas que estão embutidas nela, então ela tem um valor mais ou menos invariante estimado em 2 μF/cm2 (a capacitância total de um pedaço de membrana é proporcional à sua área). A condutância de uma bicamada lipídica pura é tão baixa, por outro lado, que em situações biológicas é sempre dominada pela condutância de vias alternativas fornecidas por moléculas incorporadas. Assim, a capacitância da membrana é mais ou menos fixa, mas a resistência é altamente variável.
A espessura de uma membrana plasmática é estimada em cerca de 7-8 nanômetros. Como a membrana é tão fina, não é necessária uma voltagem transmembrana muito grande para criar um forte campo elétrico dentro dela. Os potenciais de membrana típicos em células animais são da ordem de 100 milivolts (ou seja, um décimo de volt), mas os cálculos mostram que isso gera um campo elétrico próximo ao máximo que a membrana pode sustentar - foi calculado que uma voltagem diferença muito maior do que 200 milivolts pode causar ruptura dielétrica, isto é, arco através da membrana.
Difusão facilitada e transporte
editar
A resistência de uma bicamada lipídica pura à passagem de íons através dela é muito alta, mas as estruturas embutidas na membrana podem aumentar muito o movimento de íons, ativa ou passivamente, por meio de mecanismos chamados de transporte facilitado e difusão facilitada. Os dois tipos de estrutura que desempenham os maiores papéis são os canais iônicos e as bombas iônicas, ambos geralmente formados a partir de conjuntos de moléculas de proteínas. Os canais iônicos fornecem passagens através das quais os íons podem se mover. Na maioria dos casos, um canal iônico é permeável apenas a tipos específicos de íons (por exemplo, sódio e potássio, mas não cloreto ou cálcio), e às vezes a permeabilidade varia dependendo da direção do movimento do íon. As bombas de íons, também conhecidas como transportadores de íons ou proteínas transportadoras, transportam ativamente tipos específicos de íons de um lado da membrana para o outro, às vezes usando energia derivada de processos metabólicos para fazê-lo.
Bombas iônicas
editar
As bombas iônicas são proteínas integrais de membrana que realizam transporte ativo, ou seja, usam energia celular (ATP) para "bombear" os íons contra seu gradiente de concentração.[6] Essas bombas iônicas captam íons de um lado da membrana (diminuindo sua concentração ali) e os liberam do outro lado (aumentando sua concentração ali).
A bomba iônica mais relevante para o potencial de ação é a bomba sódio-potássio, que transporta três íons sódio para fora da célula e dois íons potássio para dentro.[7] Como consequência, a concentração de íons potássio K+ dentro do neurônio é aproximadamente 20 vezes maior que a concentração externa, enquanto a concentração externa de sódio é aproximadamente nove vezes maior que a concentração interna.[8][9] De maneira semelhante, outros íons apresentam concentrações diferentes dentro e fora do neurônio, como cálcio, cloreto e magnésio.[9]
Se os números de cada tipo de íon fossem iguais, a bomba de sódio-potássio seria eletricamente neutra, mas, devido à troca de três por dois, ela fornece um movimento líquido de uma carga positiva do intracelular para o extracelular para cada ciclo, contribuindo assim para uma diferença de tensão positiva. A bomba tem três efeitos: (1) aumenta a concentração de sódio no espaço extracelular e baixa no espaço intracelular; (2) torna a concentração de potássio alta no espaço intracelular e baixa no espaço extracelular; (3) dá ao espaço intracelular uma voltagem negativa em relação ao espaço extracelular.
A bomba de sódio-potássio é relativamente lenta em operação. Se uma célula fosse inicializada com concentrações iguais de sódio e potássio em todos os lugares, levaria horas para a bomba estabelecer o equilíbrio. A bomba opera constantemente, mas torna-se progressivamente menos eficiente à medida que as concentrações de sódio e potássio disponíveis para bombeamento são reduzidas.
As bombas de íons influenciam o potencial de ação apenas estabelecendo a proporção relativa das concentrações de íons intracelulares e extracelulares. O potencial de ação envolve principalmente a abertura e fechamento de canais iônicos e não bombas iônicas. Se as bombas de íons forem desligadas removendo sua fonte de energia ou adicionando um inibidor como a ouabaína, o axônio ainda pode disparar centenas de milhares de potenciais de ação antes que suas amplitudes comecem a decair significativamente.[6] Em particular, as bombas iônicas não desempenham nenhum papel significativo na repolarização da membrana após um potencial de ação.
Outra bomba de íons funcionalmente importante é o trocador de sódio-cálcio. Essa bomba opera de maneira conceitualmente semelhante à bomba sódio-potássio, exceto que em cada ciclo ela troca três Na+ do espaço extracelular por um Ca++ do espaço intracelular. Como o fluxo líquido de carga é para dentro, essa bomba funciona "descendo" e, portanto, não requer nenhuma fonte de energia, exceto a tensão da membrana. Seu efeito mais importante é bombear cálcio para fora – também permite um fluxo de sódio para dentro, neutralizando assim a bomba sódio-potássio, mas, como as concentrações gerais de sódio e potássio são muito mais altas que as concentrações de cálcio, esse efeito é relativamente sem importância. O resultado final do trocador sódio-cálcio é que, no estado de repouso, as concentrações de cálcio intracelular tornam-se muito baixas.
Canais iônicos
editarOs canais iônicos são proteínas integrais de membrana com um poro através do qual os íons podem viajar entre o espaço extracelular e o interior da célula. A maioria dos canais são específicos (seletivos) para um íon; por exemplo, a maioria dos canais de potássio é caracterizada por uma razão de seletividade de 1000:1 para potássio em relação ao sódio, embora os íons potássio e sódio tenham a mesma carga e diferem apenas ligeiramente em seu raio. O poro do canal é tipicamente tão pequeno que os íons devem passar por ele em ordem de arquivo único.[10][11] Os poros do canal podem ser abertos ou fechados para a passagem de íons, embora vários canais demonstrem vários níveis de subcondutância. Quando um canal está aberto, os íons permeiam através do poro do canal a favor do gradiente de concentração transmembrana para aquele íon em particular. A taxa de fluxo iônico através do canal, ou seja, amplitude de corrente de canal único, é determinada pela condutância máxima do canal e pela força eletroquímica para aquele íon, que é a diferença entre o valor instantâneo do potencial de membrana e o valor do potencial de reversão.

Um canal pode ter vários estados diferentes (correspondendo a diferentes conformações da proteína), mas cada um desses estados é aberto ou fechado. Em geral, os estados fechados correspondem a uma contração do poro – tornando-o intransponível ao íon – ou a uma parte separada da proteína, fechando o poro. Por exemplo, o canal de sódio dependente de voltagem sofre inativação, na qual uma porção da proteína oscila para dentro do poro, selando-o.[12] Essa inativação desliga a corrente de sódio e desempenha um papel crítico no potencial de ação.
Os canais iônicos podem ser classificados pela forma como respondem ao seu ambiente.[13] Por exemplo, os canais iônicos envolvidos no potencial de ação são canais sensíveis à voltagem; eles abrem e fecham em resposta à voltagem através da membrana. Canais controlados por ligantes formam outra classe importante; esses canais iônicos abrem e fecham em resposta à ligação de uma molécula de ligante, como um neurotransmissor. Outros canais iônicos abrem e fecham com forças mecânicas. Ainda outros canais iônicos - como os dos neurônios sensoriais - abrem e fecham em resposta a outros estímulos, como luz, temperatura ou pressão.
Canais de vazamento
editarOs canais de vazamento são o tipo mais simples de canal iônico, pois sua permeabilidade é mais ou menos constante. Os tipos de canais de vazamento que têm maior significado nos neurônios são os canais de potássio e cloreto. Mesmo estes não são perfeitamente constantes em suas propriedades: primeiro, a maioria deles é dependente da tensão no sentido de que eles conduzem melhor em uma direção do que na outra (em outras palavras, eles são retificadores); segundo, alguns deles são capazes de serem desligados por ligantes químicos, embora não necessitem de ligantes para operar.
Canais controlados por ligantes
editar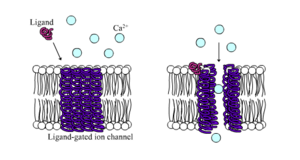
Canais iônicos controlados por ligantes são canais cuja permeabilidade é muito aumentada quando algum tipo de ligante químico se liga à estrutura da proteína. As células animais contêm centenas, senão milhares, desses tipos. Um grande subconjunto funciona como receptores de neurotransmissores - eles ocorrem em locais pós-sinápticos, e o ligante químico que os controla é liberado pelo terminal axônico pré-sináptico. Um exemplo desse tipo é o receptor AMPA, um receptor para o neurotransmissor glutamato que, quando ativado, permite a passagem de íons sódio e potássio. Outro exemplo é o receptor GABAA, um receptor para o neurotransmissor GABA que, quando ativado, permite a passagem de íons cloreto.
Os receptores de neurotransmissores são ativados por ligantes que aparecem na área extracelular, mas existem outros tipos de canais controlados por ligantes que são controlados por interações no lado intracelular.
Canais dependentes de voltagem
editarCanais iônicos dependentes de voltagem são canais cuja permeabilidade é influenciada pelo potencial de membrana. Eles formam outro grupo muito grande, com cada membro tendo uma seletividade iônica particular e uma dependência de voltagem particular. Muitos também são dependentes do tempo - em outras palavras, eles não respondem imediatamente a uma mudança de tensão, mas apenas após um atraso.
Um dos membros mais importantes desse grupo é um tipo de canal de sódio dependente de voltagem que está subjacente aos potenciais de ação. O canal é fechado no nível de voltagem de repouso, mas abre abruptamente quando a voltagem ultrapassa um certo limiar, permitindo um grande influxo de íons sódio que produz uma mudança muito rápida no potencial de membrana. A recuperação de um potencial de ação é parcialmente dependente de um tipo de canal de potássio dependente de voltagem que é fechado no nível de voltagem de repouso, mas abre como consequência da grande mudança de voltagem produzida durante o potencial de ação.
Potencial de reversão
editarO potencial de reversão (ou potencial de equilíbrio) de um íon é o valor da voltagem transmembrana no qual as forças difusivas e elétricas se contrabalançam, de modo que não há fluxo iônico líquido através da membrana. Isso significa que a voltagem transmembrana se opõe exatamente à força de difusão do íon, de modo que a corrente líquida do íon através da membrana é zero e imutável. O potencial de reversão é importante porque fornece a voltagem que atua nos canais permeáveis a esse íon, ou seja, fornece a voltagem que o gradiente de concentração do íon gera quando atua como uma bateria.
O potencial de equilíbrio de um íon particular é usualmente designado pela notação Eion.O potencial de equilíbrio para qualquer íon pode ser calculado usando a Equação de Nerst.[3] Por exemplo, o potencial de reversão para íons potássio será o seguinte:
![{\displaystyle E_{eq,K^{+}}={\frac {RT}{zF}}\ln {\frac {[K^{+}]_{o}}{[K^{+}]_{i}}},}](https://wikimedia.org/api/rest_v1/media/math/render/svg/b7eb1afcc5d7a5032ff24be5c403b43b1307b01e)
onde
- Eeq,K+ é o potencial de equilíbrio do potássio, em volts
- R é a constante universal dos gases perfeitos, equivalente a 8.314 joules·K−1·mol−1
- T é a temperatura absoluta, medida em kelvins (= K = graus Celsius + 273.15)
- z é o número de cargas elementares do íon em questão
- F é a constante de Faraday , equivalente a 96,485 coulombs·mol−1 ou J·V−1·mol−1
- [K+]o é a concentração extracelular de potássio, medida em mol·m−3 ou mmol·l−1
- [K+]i é a concentração intracelular de potássio
Mesmo que dois íons diferentes tenham a mesma carga (ou seja, K+ e Na+), eles ainda podem ter potenciais de equilíbrio muito diferentes, desde que suas concentrações externas e/ou internas sejam diferentes. Tomemos, por exemplo, os potenciais de equilíbrio de potássio e sódio nos neurônios. O potencial de equilíbrio de potássio EK é −84 mV com 5 mM de potássio externamente e 140 mM internamente. Por outro lado, o potencial de equilíbrio de sódio, ENa, é de aproximadamente +66 mV com aproximadamente 12 mM de sódio dentro e 140 mM fora.[nota 1]
Alterações no potencial de membrana durante o desenvolvimento
editarO potencial de membrana em repouso de um neurônio realmente muda durante o desenvolvimento de um organismo. Para que um neurônio eventualmente adote sua função adulta completa, seu potencial deve ser rigidamente regulado durante o desenvolvimento. À medida que um organismo progride através do desenvolvimento, o potencial de repouso da membrana torna-se mais negativo.[14] As células da glia também estão se diferenciando e proliferando à medida que o desenvolvimento progride no cérebro.[15] A adição dessas células gliais aumenta a capacidade do organismo de regular o potássio extracelular. A queda no potássio extracelular pode levar a uma diminuição do potencial de membrana de 35 mV.[16]
Excitabilidade celular
editarA excitabilidade celular é a mudança no potencial de membrana que é necessária para as respostas celulares em vários tecidos. A excitabilidade celular é uma propriedade que é induzida durante a embriogênese precoce.[17] A excitabilidade de uma célula também foi definida como a facilidade com que uma resposta pode ser desencadeada.[18] Os potenciais de repouso e limiar formam a base da excitabilidade celular e esses processos são fundamentais para a geração de potenciais graduados e de ação.
Os reguladores mais importantes da excitabilidade celular são as concentrações de eletrólitos extracelulares (ou seja, Na+, K+, Ca2+, Cl−, Mg2+ ) e proteínas associadas. Proteínas importantes que regulam a excitabilidade celular são os canais iônicos dependentes de voltagem, os transportadores iônicos, receptores de membrana e canais controlados por nucleotídeos cíclicos ativados por hiperpolarização.[19] Por exemplo, os canais de potássio e os receptores sensíveis ao cálcio são importantes reguladores da excitabilidade nos neurônios, miócitos cardíacos e muitas outras células excitáveis, como os astrócitos. [20] O íon cálcio também é o segundo mensageiro mais importante na sinalização de células excitáveis. A ativação de receptores sinápticos inicia mudanças duradouras na excitabilidade neuronal. [21] Hormônios da tireóide, suprarrenais e outros também regulam a excitabilidade celular; por exemplo, a progesterona e o estrogênio modulam a excitabilidade das células do músculo liso miometrial.
Muitos tipos celulares são considerados como tendo uma membrana excitável. As células excitáveis são neurônios, miócitos (cardíacos, esqueléticos, lisos), células endoteliais vasculares, pericitos, células justaglomerulares, células intersticiais de Cajal, muitos tipos de células epiteliais, células gliais (por exemplo, astrócitos), células mecanorreceptoras, células quimiorreceptoras, algumas células vegetais e possivelmente leucócitos. [22] Os astrócitos apresentam uma forma de excitabilidade não elétrica baseada em variações de cálcio intracelular relacionadas à expressão de vários receptores através dos quais podem detectar o sinal sináptico. Nos neurônios, existem diferentes propriedades de membrana em algumas porções da célula, por exemplo, a excitabilidade dendrítica confere aos neurônios a capacidade de detecção de coincidência de entradas espacialmente separadas.[23]
Circuito equivalente
editar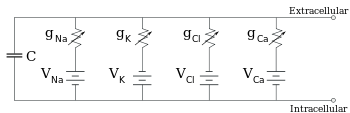
Os eletrofisiologistas modelam os efeitos das diferenças de concentração iônica, canais iônicos e capacitância da membrana em termos de um circuito equivalente, que se destina a representar as propriedades elétricas de um pequeno pedaço de membrana. O circuito equivalente consiste em um capacitor em paralelo com quatro caminhos cada um consistindo de uma bateria em série com uma condutância variável. A capacitância é determinada pelas propriedades da bicamada lipídica e é considerada fixa. Cada uma das quatro vias paralelas vem de um dos principais íons, sódio, potássio, cloreto e cálcio. A voltagem de cada via iônica é determinada pelas concentrações do íon em cada lado da membrana. A condutância de cada via iônica em qualquer ponto no tempo é determinada pelos estados de todos os canais iônicos que são potencialmente permeáveis a esse íon, incluindo canais de vazamento, canais dependentes de ligantes e canais iônicos dependentes de voltagem.
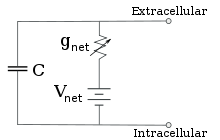
Para concentrações de íons fixas e valores fixos de condutância do canal de íons, o circuito equivalente pode ser ainda mais reduzido, usando a equação de Goldman conforme descrito abaixo, para um circuito contendo uma capacitância em paralelo com uma bateria e condutância. Em termos elétricos, este é um tipo de circuito RC (circuito de resistência-capacitância), e suas propriedades elétricas são muito simples. A partir de qualquer estado inicial, a corrente que flui através da condutância ou da capacitância decai com um curso de tempo exponencial, com uma constante de tempo de τ = RC, onde C é a capacitância do pedaço de membrana, e R = 1/gnet é a resistância líquida. Para situações realistas, a constante de tempo geralmente fica na faixa de 1 a 100 milissegundos. Na maioria dos casos, as mudanças na condutância dos canais iônicos ocorrem em uma escala de tempo mais rápida, então um circuito RC não é uma boa aproximação; no entanto, a equação diferencial usada para modelar um patch de membrana é geralmente uma versão modificada da equação do circuito RC.
Potencial de repouso
editarQuando o potencial de membrana de uma célula passa por um longo período de tempo sem mudanças significativas, ele é chamado de potencial de repouso. Este termo é usado para o potencial de membrana de células não-excitáveis, mas também para o potencial de membrana de células excitáveis na ausência de excitação. Em células excitáveis, os outros estados possíveis são potenciais de membrana graduados (de amplitude variável) e potenciais de ação. As células excitáveis incluem neurônios, células musculares e algumas células secretoras nas glândulas. Mesmo em outros tipos de células, no entanto, a voltagem da membrana pode sofrer alterações em resposta a estímulos ambientais ou intracelulares. Por exemplo, a despolarização da membrana plasmática parece ser um passo importante na morte celular programada.[24]
As interações que geram o potencial de repouso são modeladas pela equação de Goldman.[3] A equação é semelhante em forma à equação de Nernst mostrada anteriormente, pois é baseada nas cargas dos íons em questão, bem como na diferença de suas concentrações internas e externas. A equação de Goldman, entretanto, também leva em consideração a permeabilidade relativa da membrana plasmática para cada íon em questão.
![{\displaystyle E_{m}={\frac {RT}{F}}\ln {\left({\frac {P_{\mathrm {K} }[\mathrm {K} ^{+}]_{\mathrm {out} }+P_{\mathrm {Na} }[\mathrm {Na} ^{+}]_{\mathrm {out} }+P_{\mathrm {Cl} }[\mathrm {Cl} ^{-}]_{\mathrm {in} }}{P_{\mathrm {K} }[\mathrm {K} ^{+}]_{\mathrm {in} }+P_{\mathrm {Na} }[\mathrm {Na} ^{+}]_{\mathrm {in} }+P_{\mathrm {Cl} }[\mathrm {Cl} ^{-}]_{\mathrm {out} }}}\right)}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/4b29151571b6fb807674bf342d01f178908a1a98)
Os três íons que aparecem nesta equação são potássio (K+), sódio (Na+) e cloreto (Cl−). O cálcio é omitido, mas pode ser adicionado para lidar com situações em que desempenha um papel significativo.[25] Sendo um ânion, o cloreto é tratado de forma diferente dos íons catiônicos; a concentração intracelular está no numerador e a concentração extracelular no denominador, o que é invertido para cátions. Pi representa a permeabilidade relativa do íon tipo i.
Em essência, a fórmula de Goldman expressa o potencial de membrana como uma média ponderada dos potenciais de reversão para os tipos de íons individuais, ponderados pela permeabilidade. (Embora o potencial de membrana mude cerca de 100 mV durante um potencial de ação, as concentrações de íons dentro e fora da célula não mudam significativamente. Eles permanecem próximos de suas respectivas concentrações quando a membrana está em potencial de repouso.) Na maioria das células animais, a permeabilidade ao potássio é muito maior no estado de repouso do que a permeabilidade ao sódio. Como consequência, o potencial de repouso é geralmente próximo do potencial de reversão do potássio.[3] A permeabilidade ao cloreto pode ser alta o suficiente para ser significativa, mas, ao contrário dos outros íons, o cloreto não é bombeado ativamente e, portanto, equilibra em um potencial de reversão muito próximo do potencial de repouso determinado pelos outros íons.
Os valores do potencial de membrana em repouso na maioria das células animais geralmente variam entre o potencial de reversão do potássio (geralmente em torno de -80 mV) e em torno de -40 mV. O potencial de repouso em células excitáveis (capazes de produzir potenciais de ação) é geralmente próximo a -60 mV – voltagens mais despolarizadas levariam à geração espontânea de potenciais de ação. Células imaturas ou indiferenciadas mostram valores altamente variáveis de voltagem de repouso, geralmente significativamente mais positivos do que em células diferenciadas.[26] Em tais células, o valor do potencial de repouso se correlaciona com o grau de diferenciação: células indiferenciadas em alguns casos podem não apresentar nenhuma diferença de voltagem transmembrana.
A manutenção do potencial de repouso pode ter um alto custo metabólico para uma célula devido à sua necessidade de bombeamento ativo de íons para neutralizar as perdas devido aos canais de vazamento. O custo é maior quando a função da célula requer um valor especialmente despolarizado da voltagem da membrana.[27] Esse potencial de membrana elevado permite que as células respondam muito rapidamente a estímulos; o custo é que a manutenção do potencial de repouso pode consumir mais de 20% do ATP celular total.[28]
Por outro lado, o alto potencial de repouso em células indiferenciadas não necessariamente incorre em um alto custo metabólico. Este aparente paradoxo é resolvido pelo exame da origem desse potencial de repouso. As células pouco diferenciadas são caracterizadas por uma resistência de entrada extremamente alta,[26] o que implica que poucos canais de vazamento estão presentes nesta fase da vida celular. Como resultado aparente, a permeabilidade do potássio torna-se semelhante à dos íons sódio, o que coloca o potencial de repouso entre os potenciais de reversão do sódio e do potássio, conforme discutido acima. As correntes de fuga reduzidas também significam que há pouca necessidade de bombeamento ativo para compensar, portanto, baixo custo metabólico.
Potenciais graduados
editarComo explicado acima, o potencial em qualquer ponto da membrana de uma célula é determinado pelas diferenças de concentração de íons entre as áreas intracelular e extracelular e pela permeabilidade da membrana a cada tipo de íon. As concentrações de íons normalmente não mudam muito rapidamente (com exceção de Ca2+, onde a concentração intracelular da linha de base é tão baixa que mesmo um pequeno influxo pode aumentá-la em ordens de magnitude), mas as permeabilidades dos íons podem mudar em uma fração de milissegundo, como resultado da ativação de canais iônicos controlados por ligantes. A mudança no potencial de membrana pode ser grande ou pequena, dependendo de quantos canais iônicos são ativados e de que tipo eles são, e pode ser longa ou curta, dependendo do tempo que os canais permanecem abertos. Alterações desse tipo são chamadas de potenciais graduados, em contraste com os potenciais de ação, que têm amplitude e curso de tempo fixos.
Como pode ser derivado da equação de Goldman mostrada acima, o efeito de aumentar a permeabilidade de uma membrana a um tipo particular de íon desloca o potencial de membrana para o potencial de reversão desse íon. Assim, a abertura de canais de Na+ desloca o potencial de membrana para o potencial de reversão de Na+, que geralmente é em cerca de +100 mV. Da mesma forma, a abertura de canais de K+ desloca o potencial de membrana para cerca de –90 mV, e a abertura de canais de Cl− o desloca para cerca de –70 mV (potencial de repouso da maioria das membranas). Dessa forma, os canais de Na+ deslocam o potencial de membrana em uma direção positiva, canais de K+ deslocam a membrana em uma direção negativa (exceto quando a membrana está hiperpolarizada em um valor mais negativo que o potencial de reversão de K+), e canais de Cl− tendem desloca-lo em direção do potencial de repouso.
Potenciais de membrana graduados são particularmente importantes em neurônios, onde são produzidos por sinapses – uma mudança temporária no potencial de membrana produzida pela ativação de uma sinapse por um único potencial graduado ou de ação é chamado de potencial pós-sináptico. Os neurotransmissores que agem para abrir os canais de Na+ normalmente fazem com que o potencial de membrana se torne mais positivo, enquanto os neurotransmissores que ativam os canais de K+ normalmente fazem com que ele se torne mais negativo; aqueles que inibem esses canais tendem a ter o efeito oposto.
Se um potencial pós-sináptico é considerado excitatório ou inibitório depende do potencial de reversão para os íons dessa corrente e do limiar para a célula disparar um potencial de ação (cerca de -50mV). Uma corrente pós-sináptica com potencial de reversão acima do limiar, como uma corrente típica de Na+, é considerada excitatória. Uma corrente com um potencial de reversão abaixo do limite, como uma corrente K+ típica, é considerada inibitória. Uma corrente com um potencial de reversão acima do potencial de repouso, mas abaixo do limiar, não irá por si só provocar potenciais de ação, mas produzirá oscilações de potencial de membrana subliminares. Assim, neurotransmissores que agem para abrir canais de Na+ produzem potenciais pós-sinápticos excitatórios, ou EPSPs, enquanto neurotransmissores que agem para abrir canais de K+ ou Cl− normalmente produzem potenciais pós-sinápticos inibitórios, ou IPSPs. Quando vários tipos de canais são abertos no mesmo período de tempo, seus potenciais pós-sinápticos se somam (são adicionados juntos).
Outros valores
editarDo ponto de vista biofísico o potencial de membrana em repouso é meramente o potencial de membrana que resulta das permeabilidades de membrana que predominam quando a célula está em repouso. A equação acima de médias ponderadas sempre se aplica, mas a abordagem a seguir pode ser mais facilmente visualizada.↵Em qualquer momento, existem dois fatores para um íon que determinam quanta influência esse íon terá sobre o potencial de membrana de uma célula:
- A força motriz do íon
- A permeabilidade do íon
Se a força motriz for alta, então o íon está sendo "empurrado" através da membrana. Se a permeabilidade for alta, será mais fácil para o íon se difundir através da membrana.
- A força motriz é a força elétrica líquida disponível para mover esse íon através da membrana. É calculado como a diferença entre a voltagem em que o íon "quer" estar (seu potencial de equilíbrio) e o potencial de membrana real (Em). Então, em termos formais, a força motriz para um íon = Em - Eion
- Por exemplo, em nosso potencial de repouso calculado anteriormente de -73 mV, a força motriz no potássio é 7 mV: (−73 mV) − (−80 mV) = 7 mV. A força motriz sobre o sódio seria (−73 mV) − (60 mV) = −133 mV.
- Permeabilidade é uma medida de quão facilmente um íon pode atravessar a membrana. Normalmente é medida como a condutância (elétrica) e a unidade, siemens, corresponde a 1 C·s−1·V−1, que é um coulomb por segundo por volt de potencial.
Assim, em uma membrana em repouso, enquanto a força motriz do potássio é baixa, sua permeabilidade é muito alta. O sódio tem uma enorme força motriz, mas quase nenhuma permeabilidade de repouso. Nesse caso, o potássio carrega cerca de 20 vezes mais corrente que o sódio e, portanto, tem 20 vezes mais influência sobre o Em do que o sódio.
No entanto, considere outro caso - o pico do potencial de ação. Aqui, a permeabilidade ao Na é alta e a permeabilidade ao K é relativamente baixa. Assim, a membrana se move para ENa e longe de EK.
Quanto mais íons são permeáveis, mais complicado se torna prever o potencial de membrana. No entanto, isso pode ser feito usando a equação de Goldman-Hodgkin-Katz ou a equação das médias ponderadas. Ao conectar os gradientes de concentração e as permeabilidades dos íons em qualquer instante no tempo, pode-se determinar o potencial de membrana naquele momento. O que as equações de GHK significam é que, a qualquer momento, o valor do potencial de membrana será uma média ponderada dos potenciais de equilíbrio de todos os íons permeantes. A "ponderação" é a permeabilidade relativa dos íons através da membrana.
Efeitos e implicações
editarEnquanto as células gastam energia para transportar íons e estabelecer um potencial transmembrana, elas usam esse potencial para transportar outros íons e metabólitos, como o açúcar. O potencial transmembranar das mitocôndrias impulsiona a produção de ATP, que é a moeda comum da energia biológica.
As células podem utilizar a energia que armazenam no potencial de repouso para conduzir potenciais de ação ou outras formas de excitação. Essas mudanças no potencial de membrana permitem a comunicação com outras células (como os potenciais de ação) ou iniciam mudanças dentro da célula, o que acontece em um óvulo quando é fertilizado por um espermatozoide.
Nas células neuronais, um potencial de ação começa com um fluxo de íons sódio para dentro da célula através dos canais de sódio, resultando em despolarização, enquanto a recuperação envolve um fluxo de potássio para fora através dos canais de potássio. Ambos os fluxos ocorrem por difusão passiva.
Ver também
editarNotes
editar- ↑ Observe que os sinais de ENa e EK são opostos. Isso ocorre porque o gradiente de concentração de potássio é direcionado para fora da célula, enquanto o gradiente de concentração de sódio é direcionado para dentro da célula. Os potenciais de membrana são definidos em relação ao exterior da célula; assim, um potencial de -70 mV implica que o interior da célula é negativo em relação ao exterior.
Referências
- ↑ Bruce, Alberts (18 de novembro de 2014). Molecular biology of the cell Sixth ed. New York, NY: [s.n.] ISBN 9780815344322. OCLC 887605755
- ↑ Abdul Kadir, Lina; Stacey, Michael; Barrett-Jolley, Richard (2018). «Emerging Roles of the Membrane Potential: Action Beyond the Action Potential». Frontiers in Physiology (em English). 9. ISSN 1664-042X. PMID 30519193. doi:10.3389/fphys.2018.01661

- ↑ a b c d e Bullock TH, Orkand R, Grinnell A (1977). Introduction to Nervous Systems. New York: W. H. Freeman. ISBN 0-7167-0030-1
- ↑ Mummert H, Gradmann D (1991). «Action potentials in Acetabularia: measurement and simulation of voltage-gated fluxes». Journal of Membrane Biology. 124 (3): 265–73. PMID 1664861. doi:10.1007/BF01994359
- ↑ «Chapter 2. Simple Diffusion across the Membrane Barrier». Transport and Diffusion across Cell Membranes. San Diego: Academic Press. 1986. pp. 69–112. ISBN 978-0-12-664661-0
- ↑ a b Hodgkin AL, Keynes RD (1955). «Active transport of cations in giant axons from Sepia and Loligo». J. Physiol. 128 (1): 28–60. PMC 1365754 . PMID 14368574. doi:10.1113/jphysiol.1955.sp005290
- ↑ Caldwell PC, Hodgkin AL, Keynes RD, Shaw TI (1960). «The effects of injecting energy-rich phosphate compounds on the active transport of ions in the giant axons of Loligo». J. Physiol. 152 (3): 561–90. PMC 1363339 . PMID 13806926. doi:10.1113/jphysiol.1960.sp006509
- ↑ Steinbach HB, Spiegelman S (1943). «The sodium and potassium balance in squid nerve axoplasm». J. Cell. Comp. Physiol. 22 (2): 187–96. doi:10.1002/jcp.1030220209
- ↑ a b Hodgkin AL (1951). «The ionic basis of electrical activity in nerve and muscle». Biol. Rev. 26 (4): 339–409. doi:10.1111/j.1469-185X.1951.tb01204.x
- ↑ Eisenman G (1961). «On the elementary atomic origin of equilibrium ionic specificity». In: A Kleinzeller; A Kotyk. Symposium on Membrane Transport and Metabolism. New York: Academic Press. pp. 163–79Eisenman G (1965). «Some elementary factors involved in specific ion permeation». Proc. 23rd Int. Congr. Physiol. Sci., Tokyo. Amsterdam: Excerta Med. Found. pp. 489–506
- ↑ Diamond JM, Wright EM (1969). «Biological membranes: the physical basis of ion and nonekectrolyte selectivity». Annual Review of Physiology. 31: 581–646. PMID 4885777. doi:10.1146/annurev.ph.31.030169.003053
- ↑ Cai SQ, Li W, Sesti F (2007). «Multiple modes of a-type potassium current regulation». Curr. Pharm. Des. 13 (31): 3178–84. PMID 18045167. doi:10.2174/138161207782341286
- ↑ Goldin AL (2007). «Neuronal Channels and Receptors». In: Waxman SG. Molecular Neurology. Burlington, MA: Elsevier Academic Press. pp. 43–58. ISBN 978-0-12-369509-3
- ↑ Sanes, Dan H.; Takács, Catherine (1 de junho de 1993). «Activity-dependent Refinement of Inhibitory Connections». European Journal of Neuroscience (em inglês). 5 (6): 570–574. ISSN 1460-9568. PMID 8261131. doi:10.1111/j.1460-9568.1993.tb00522.x
- ↑ KOFUJI, P.; NEWMAN, E. A. (1 de janeiro de 2004). «Potassium buffering in the central nervous system». Neuroscience. 129 (4): 1045–1056. ISSN 0306-4522. PMC 2322935 . PMID 15561419. doi:10.1016/j.neuroscience.2004.06.008
- ↑ Sanes, Dan H.; Reh, Thomas A (1 de janeiro de 2012). Development of the nervous system Third ed. [S.l.]: Elsevier Academic Press. pp. 211–214. ISBN 9780080923208. OCLC 762720374
- ↑ Tosti, Elisabetta (28 de junho de 2010). «Dynamic roles of ion currents in early development». Molecular Reproduction and Development. 77 (10): 856–867. ISSN 1040-452X. PMID 20586098. doi:10.1002/mrd.21215
- ↑ Boyet, M.R.; Jewell, B.R. (1981). «Analysis of the effects of changes in rate and rhythm upon electrical activity in the heart». Progress in Biophysics and Molecular Biology. 36 (1): 1–52. ISSN 0079-6107. PMID 7001542. doi:10.1016/0079-6107(81)90003-1
- ↑ Spinelli, Valentina; Sartiani, Laura; Mugelli, Alessandro; Romanelli, Maria Novella; Cerbai, Elisabetta (2018). «Hyperpolarization-activated cyclic-nucleotide-gated channels: pathophysiological, developmental, and pharmacological insights into their function in cellular excitability». Canadian Journal of Physiology and Pharmacology. 96 (10): 977–984. ISSN 0008-4212. PMID 29969572. doi:10.1139/cjpp-2018-0115
- ↑ Jones, Brian L.; Smith, Stephen M. (30 de março de 2016). «Calcium-Sensing Receptor: A Key Target for Extracellular Calcium Signaling in Neurons». Frontiers in Physiology. 7: 116. ISSN 1664-042X. PMC 4811949 . PMID 27065884. doi:10.3389/fphys.2016.00116
- ↑ Debanne, Dominique; Inglebert, Yanis; Russier, Michaël (2019). «Plasticity of intrinsic neuronal excitability» (PDF). Current Opinion in Neurobiology (em inglês). 54: 73–82. PMID 30243042. doi:10.1016/j.conb.2018.09.001
- ↑ Davenport, Bennett; Li, Yuan; Heizer, Justin W.; Schmitz, Carsten; Perraud, Anne-Laure (23 de julho de 2015). «Signature Channels of Excitability no More: L-Type Channels in Immune Cells». Frontiers in Immunology. 6: 375. ISSN 1664-3224. PMC 4512153 . PMID 26257741. doi:10.3389/fimmu.2015.00375
- ↑ Sakmann, Bert (21 de abril de 2017). «From single cells and single columns to cortical networks: dendritic excitability, coincidence detection and synaptic transmission in brain slices and brains». Experimental Physiology. 102 (5): 489–521. ISSN 0958-0670. PMC 5435930 . PMID 28139019. doi:10.1113/ep085776
- ↑ Franco R, Bortner CD, Cidlowski JA (2006). «Potential roles of electrogenic ion transport and plasma membrane depolarization in apoptosis». J. Membr. Biol. 209 (1): 43–58. PMID 16685600. doi:10.1007/s00232-005-0837-5
- ↑ Spangler SG (1972). «Expansion of the constant field equation to include both divalent and monovalent ions». Alabama Journal of Medical Sciences. 9 (2): 218–23. PMID 5045041
- ↑ a b Magnuson DS, Morassutti DJ, Staines WA, McBurney MW, Marshall KC (1995). «In vivo electrophysiological maturation of neurons derived from a multipotent precursor (embryonal carcinoma) cell line». Developmental Brain Research. 84 (1): 130–41. PMID 7720212. doi:10.1016/0165-3806(94)00166-W
- ↑ Juusola M, Kouvalainen E, Järvilehto M, Weckström M (1994). «Contrast gain, signal-to-noise ratio, and linearity in light-adapted blowfly photoreceptors». J Gen Physiol. 104 (3): 593–621. PMC 2229225 . PMID 7807062. doi:10.1085/jgp.104.3.593
- ↑ Laughlin SB, de Ruyter van Steveninck RR, Anderson JC (1998). «The metabolic cost of neural information». Nat. Neurosci. 1 (1): 36–41. PMID 10195106. doi:10.1038/236